

Catálogo de dinosaurios argentinos: los 50 hallazgos del tercer milenio

1 de oct. de 2025 — Luis Paz

En la primera parte de este Catálogo de Dinosaurios Argentinos repasamos los antecedentes de la paleontología local en manos extranjeras, vimos cómo se volvió casi una causa nacional en la segunda mitad del siglo XX y recordamos los principales descubrimientos de dinos argentos hasta 1999. En esta segunda parte vamos a ver qué pasó desde el cambio de milenio.
A partir del 2000, los hitos de la paleontología argentina explotaron por una combinación de factores científicos, legales y logísticos. Por un lado, la profesionalización y expansión de los equipos locales (Conicet, museos, universidades) con campañas, laboratorios, colecciones y proyectos como el del Centro Paleontológico Lago Barreales, de la Universidad Nacional del Comahue. Esto fue posible también por la consolidación de un marco legal nacional que ordenó permisos, colecciones y la lucha contra el tráfico (con la Ley 25.743, de 2003), y que puso al Museo Argentino de Ciencias Naturales (MACN) como autoridad de aplicación. Y, por supuesto, también mejoraron las herramientas y tecnologías de cartografía de detalle, geoquímica o magnetoestratigrafía. E incluso los desarrollos en logística e infraestructura nacional abrieron cortes nuevos que dieron paso a descubrimientos: por caso, el Ekrixinatosaurus encontrado gracias a una obra de gasoducto.
En este ritmo acelerado de descubrimientos resultó crucial el rol de las instituciones públicas. Fundamentalmente el tándem MACN/Conicet con sus colecciones y autoridad legal; los museos provinciales y municipales con sus depósitos y exhibiciones; y las universidades nacionales con sus laboratorios de campo y equipos de investigadores. Todas instituciones financiadas por alguna escala del Estado (nacional, provincial o municipal).

La tormenta perfecta
Según la información disponible, son al menos 50 los dinosaurios que habitaron suelo prehistórico argentino cuyos restos fueron encontrados en los últimos 25 años. El número es otra coronación de gloria, por supuesto, pero además de las condiciones para que los bichos más majestuosos de la prehistoria hayan vivido acá, lo que llama la atención también es la forma en la que esos restos se conservaron, ofreciendo en muchos casos casi que esqueletos completos.
Hay una clave geológica para esto que es la Cuenca Neuquina, un sistema retroarco/antepaís con subsidencia sostenida durante el Mesozoico, en el que se acumularon miles de metros de areniscas, limos y tobas del Grupo Neuquén (Cretácico), en ríos entrelazados, planicies de inundación y márgenes de desierto eólico (Candeleros, Huincul, Portezuelo, Plottier, Bajo de la Carpa, Anacleto). Ese apilamiento registra ecosistemas muy distintos (riberas boscosas, dunas, lagunas efímeras), lo que multiplica nichos y probabilidad de enterramiento. Por eso la cuenca es una de las más completas en vertebrados mesozoicos del mundo, y nutre gran parte de la diversidad patagónica.
De esa sopa primitiva o tormenta perfecta para el esplendor sáurico no se termina de desprender, de todos modos, la explicación de cómo y por qué se conservan tan bien los restos. Y en eso aparecen dos motivos repetidos: el rápido enterramiento (por ejemplo al norte de Río Negro, donde La Buitrera tenía donas gigantescas que enterraban organismos) y el volcanismo (que ayuda a la mineralización temprana, como en la zona de Auca Mahuevo, de la Formación Anacleto). Contextos que maximizan los detalles anatómicos y las trazas de los fósiles.
Por otro lado, aunque los dinosaurios vivieron bajo climas mesozoicos variados, la Patagonia moderna es árida: poca vegetación, suelos sueltos y erosión eólica/fluvial continua dejan aflorar grandes ventanas cretácicas, de Neuquén a Santa Cruz. Esa aridez facilita la detección de huesos "en superficie", lo que no sucede en selvas densas.
Ahora sí, sin mucho más, los 50 dinosaurios identificados en Argentina en lo que va del siglo XXI, incluyendo el reciente Joaquinraptor casali. Y son tantos que vamos a tener que hacer una clasificación por alimentación.
THEROPODA (carnívoros y afines)
Son un clado de dinosaurios caracterizados por ser mayoritariamente carnívoros y caminar sobre dos patas (bípedos obligados).
ABELISÁURIDOS
Eran los carnívoros dominantes del Cretácico del Hemisferio Sur. Se caracterizaban por tener brazos extremadamente reducidos y ornamentaciones en la cabeza como cuernos, crestas y protuberancias. El grupo al que pertenece Carnotaurus sastrei.
Ilokelesia aguadagrandensis (1998)
Abelisáurido de la Formación Huincul, Plaza Huincul (Neuquén), edad estimada entre 98-90 Ma. Descrito por Coria y Salgado; de 5 m de largo y estimado de 800 kg de peso. El nombre proviene del mapuche ilo que es "carne" y kelesio "lagarto", y de la localidad en la que lo encontraron Aguada Grande. Presenta rasgos transicionales en cráneo y cintura pélvica que ayudan a anclar caracteres primitivos del clado. Su material contribuyó a refinar las características que definen a la familia y entender cómo evoluciono la apariencia de los abelisáuridos patagónicos.
Aucasaurus garridoi (2002)
Su nombre significa "lagarto de Auca de Alberto Garrido" por la zona en la que fue encontrado, Auca Mahuevo (Neuquén), de la Formación Anacleto (Santoniano–Campaniano, ~ 75 Ma). Abelisáurido mediano de 7 m de largo y aproximadamente 700 kg. Hallado por equipos del Museo Carmen Funes/Proyecto Auca Mahuevo y descrito por Coria, Chiappe y Dingus. Comparado con Carnotaurus, tiene brazos y el cráneo más largos, y poseía 2 pequeñas protuberancias que no llegaban a cuernos. Contexto con abundantes nidadas de titanosaurios y niveles tobáceos que favorecieron la preservación. Importante para la diversidad y morfología craneal de abelisáuridos patagónicos.
Ekrixinatosaurus novasi (2004)
Sus restos provienen de de la Formación Candeleros (Cuenca Neuquina ~ 98 - 97 Ma). Su nombre significa "lagarto nacido de la explosión de Novas" debido a que fue descubierto luego de una explosión por operaciones mineras; descrito por Calvo y colegas. Abelisáurido de gran tamaño de 10 m de largo con un cráneo proporcionalmente más grande que otros abelisáuridos. Representa una fase temprana de la radiación abelisáurida en el Hemisferio Sur, convivio con otros grandes dinosaurios carnívoros como Giganotosaurus. Su contexto de hallazgo subraya el rol de la infraestructura moderna en la prospección paleontológica.
Skorpiovenator bustingorryi (2009)
Abelisáurido de la Formación Huincul (Neuquén ~ 95-93 Ma). Descrito por Canale y colaboradores, con esqueleto articulado muy bien preservado. Su nombre significa "cazador de escorpiones" alude a los escorpiones locales; y el epíteto especifico honra a Manuel Bustingorry, dueño del campo donde fue encontrado. De un largo total de 6 m, el cráneo era corto y robusto, ornamentado con crestas, tubérculos y nódulos. Es clave para caracterizar la variación craneal y la sistemática interna de Abelisauridae en el Cretácico medio–tardío.
Tralkasaurus cuyi (2019)
Dinosaurio de la Formación Huincul (Río Negro)(97-93 Ma), descrito por Cerroni y colaboradores. Abelisáurido pequeño–mediano de 6 m de largo. El nombre proviene del mapudungun "tralka" que significa "trueno" y al departamento El Cuy. Cráneo robusto y proporciones que sugieren partición de nicho respecto de abelisáuridos mayores. Posee un conjunto de características que no permiten del todo entender si es un abelisáurido primitivo o está dentro de los más especializados.
Niebla antiqua (2020)
Abelisáurido de la Formación Allen (Río Negro) (72-66 Ma), descrito por Aranciaga Rolando y colaboradores. El nombre hace referencia a los días neblinosos durante su excavación. Era relativamente pequeño de 4 a 5 m de largo. El material es fragmentario y no articulado. Esta dentro del grupo más especializado de abelisáuridos y poseía una estructuración de los huesos pectorales única dentro del grupo.
Llukalkan aliocranianus (2021)
Abelisáurido de la Formación Bajo de la Carpa (Neuquén/Río Negro) (86,3 - 83,6 Ma). Descrito por Gianechini y colegas. Su nombre Llukalkan viene del mapuche y significa "el que causa miedo", y del latín aliocranianus, "con cráneo diferente". Cráneo con cavidades del oído medio y neumatización peculiares; posible sensibilidad auditiva diferenciada. Coexistió con otros abelisáuridos, indicando alta riqueza local del grupo en ambientes santonianos semiáridos del Grupo Neuquén.
Elemgasem nubilus (2022)
Abelisáurido pequeño de la Formación Portezuelo (Neuquén) (90,5 - 88,5 Ma), descrito por Baiano y colaboradores. El nombre Elemgasem es el de un dios tehuelche dueño de los animales capaz de petrificar a los demás, y nubilus hace referencia a los días nubosos de su excavación. El fósil presenta espondiloartropatía (enfermedad de las articulaciones vertebrales) el cual lo convierte en el primer caso de dinosaurio no aviano con esa enfermedad.
Koleken inakayali (2023)
Abelisaurio de la Formación La Colonia (Chubut) (69 Ma), descrito por Pol y colaboradores. El espécimen corresponde a un subadulto de 6 años de edad y de 5 m de largo. El nombre kolenken proviene del teushen y significa "el que proviene de la arcilla y el agua", ya que fue encontrado en sedimentos correspondientes a barro arcilloso de un ambiente estuariano. El nombre específico inakayali honra al ultimo jefe tehuelche, Inayakal. Se ha planteado que podría ser un estado juvenil de Carnotaurus, ya que presenta muy pocas diferencias con este, sobre todo la falta de cuernos en el cráneo la cual podría explicarse como parte del crecimiento de animal si fueran de la misma especie.
CARCARODONTOSÁURIDOS
Eran los predadores más grandes del hemisferio Sur que vivieron entre el Jurásico superior (154 Ma) y el Cretácico superior (89,3 Ma). Eran dinosaurios de gran longitud y tamaño pero gráciles en comparación con los tiranosáuridos. Presentaban dientes delgados y aserrados. Dentro de este grupo se encuentra Giganotosaurus.
Tyrannotitan chubutensis (2005)
Carcarodontosáurido de Chubut (Formación Cerro Barcino)(118-112 Ma). Descrito por Novas y colaboradores. Su nombre significa "titán tirano de Chubut". De un largo de 12 m de largo y un estimado de 7 toneladas es el carcarodontosaurio más antiguo conocido. Cráneo largo con dientes aserrados de gran tamaño, robustas vértebras y extremidades poderosas; depredador ápice previo al auge de megaraptoridos. Su talla y anatomía lo posicionan entre los mayores terópodos gondwánicos, se caracteriza por la falta de neumaticidad (bolsas de aire en los huesos que alivianan la estructura) en la zona sacra y vertebras de la cola.

Mapusaurus roseae (2006)
Carcharodontosáurido de la Formación Huincul (Neuquén) (97 - 93,3 Ma), descrito por Coria y Currie, encontrándose entre 7 y 9 individuos en la excavación. Se estima medía 12 m de largo y pesaba 6 toneladas. El nombre proviene del mapuche Mapu que significa "tierra", y roseae por el color de las rocas en las que lo encontraron, es decir "reptil de la tierra rosacea". No se ha determinado si la acumulación de esqueletos sugiere posible gregarismo o es el resultado de una trampa natural.
MEGARAPTÓRIDOS
Dinosaurios terópodos que se caracterizan por presentan brazos largos y robustos con poderosas garras en forma de hoz. Fueron los predadores de mayor tamaño del Hemisferio Sur durante el Cretácico (entre los 128 - 66 Ma). En este grupo definido por la especie Megaraptor namunhuaiquii.
Aerosteon riocoloradensis (2008)
Megaraptórido de la Formación Río Colorado (Mendoza) con neumatización extrema del esqueleto; descrito por Sereno y colegas. Su nombre proviene del griego aeros y osteon que significa "huesos aireados" por la presencia de sus huesos "huecos" (neumaticidad), estructura que presentan las aves también. Evidencia no solo la relación estrecha de terópodos-aves, sino que también aporta dato sobre la posible anatomía interna que presentaban los dinosaurios.
Murusraptor barrosaensis (2016)
Megaraptórido de la Formación Sierra Barrosa (Neuquén) (93 - 89 Ma), descrito por Coria y Currie. Su nombre significa "ladrón de la pared" ya que el espécimen fue encontrado en la pared de un cañón. De aproximadamente 6,5 m de largo, presenta garras manuales hipertrofiadas, caja torácica amplia y rasgos axiales diagnósticos del clado. Es un ejemplar clave para definir las características que hacen distintivos a los Megaraptoridae y su relación con otros tetanuros. Además el espécimen presenta signos de infecciones en el lado izquierdo de la cabeza que deformaron el cráneo, se cree que fueron provocadas por otro terópodo.
Aoniraptor libertatem (2016)
Megaraptor de la Formación Huincul (Río Negro/Neuquén)(97 - 95 Ma) descrito por Motta y colaboradores. Su nombre es una combinación de tehuelche aoni que significa "sur" y del latín raptor que es "ladrón", libertatem hace referencia a la declaración de la independencia del país que cumplía 200 años cuando se describió a este dinosaurio en 2010. De aproximadamente 6 m de largo.
Maip macrothorax (2022)
Megaraptórido gigante (de 9 m de largo) de la Formación Chorrillo (Santa Cruz) (70 Ma). Descrito por Aranciaga Rolando y colegas. Maip es el nombre de un ser mitológico de Ainokenk, un ser malicioso conocido como "la sombra de la muerte que mata con viento frío" ya que fue extraído a 30 km del Calafate en una zona de altura. El nombre específico macrothorax o "pecho grande" hace referencia a la amplia caja toráxica de este dinosaurio. Es el megaraptórido más grande de América del Sur, indicando como este grupo alcanzo tallas más grandes hacia finales del Cretácico.
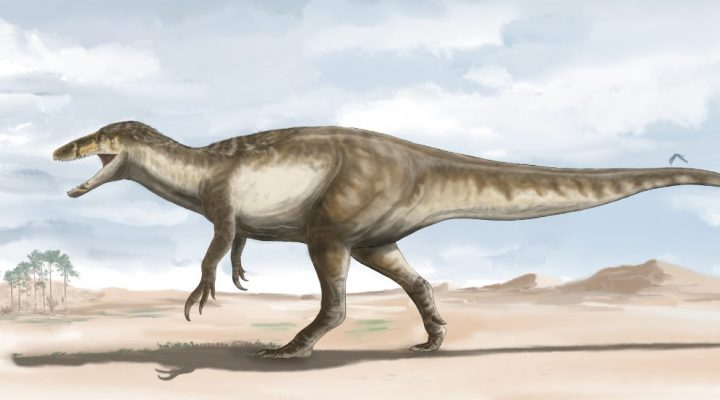
Joaquinraptor casali (2025)
Megaraptórido grande de la Formación Lago Colhué Huapi (Chubut) (70 - 66 Ma). Descrito por Ibiricu y colegas. Con un estimado de casi 7 m de largo y un peso de 700 kg se infiere que el espécimen tenia 19 años de edad. Su nombre genérico hace honor a Joaquín el primer hijo del autor y el nombre informal del paraje donde se lo encontró "Valle Joaquín", mientras que el epíteto específico hace referencia a Dr. Gabriel A. Casal. El holotipo fue encontrado con huesos de la pata de un cocodrilomorfo atrapado en la mandíbula: rara evidencia directa de dieta.
RAPTORES UNENLAGINOS
Dinosaurios raptores pequeños y medianos, probablemente emplumados de cuerpos estilizados y ágiles. Los unenlaginos son exclusivamente sudamericanos que vivieron a final del Cretácico, que se caracterizan por tener un hocico alargado y son los parientes más cercanos a las aves dentro de los raptores.
Buitreraptor gonzalezorum (2005)
Unenlagino de La Buitrera, Formación Candeleros (Río Negro) (98 - 97 Ma). Descrito por Makovicky, Apesteguía y Agnolín. Su nombre significa "rapaz de la Buitrera de Fabián Gonzáles y Jorge González". Se calcula que medía 1,2 m de largo y llego a pesar 5 kg. Es el espécimen más completo de dromeosáurido encontrado en el Hemisferio Sur. Posiblemente cazaba pequeños animales como mamíferos y pequeños lagartos.
Austroraptor cabazai (2008)
Unenlagino de gran tamaño (≈5 m) de la Formación Allen (Río Negro) (71 - 70 Ma). Descrito por Novas y colaboradores, su nombre significa "ladrón/raptor del sur". Se estima alcazaba los 6 m de largo y pesaba 350 kg, lo que lo convierte en uno de los raptores más grandes conocidos. Posee un cráneo elongado con dientes pequeños cónicos sugiere alimentación piscívora. Es el registro más joven para los dromeosáuridos (raptores), y el más grande, del hemisferio sur.
Pamparaptor micros (2011)
Pequeño unenlagino de la Formación Portezuelo (Neuquén) (93,3 - 85,5 Ma), descrito por Porfiri, Calvo y Santos. Su nombre significa "raptor de las pampas pequeño". El espécimen consta de el miembro del pie articulado. Se cree que el tamaño total era de entre 0,5 a 0,7 m de largo.
ENIGMAS EVOLUTIVOS
Aquí agrupe algunos dinosaurios cuyas relaciones con otros grupos no están claras o se siguen debatiendo o incluso no tienen una ubicación específica aun en el contexto evolutivo de los terópodos.
Bicentenaria argentina (2012)
Coelurosaurio basal de la Formación Candeleros (Río Negro) (~ 90 Ma), descrito por Novas y colegas. De un tamaño estimado de 2,5 a 3 m de largo y un peso de 40 kg. El nombre es en honor a los 200 años de independencia de la República Argentina. En la excavación se encontraron múltiples individuos, predominantemente juveniles; debido a que había varios individuos de distintas edades los autores interpretaron que era una pequeña manada que pereció por una inundación.
Alnashetri cerropoliciensis (2012)
Pequeño terópodo de La Buitrera, Formación Candeleros (Río Negro) (95 Ma). Descrito por Makovicky y colaboradores. Esta dentro de un grupo llamado Alvarezsauroidea que se caracterizan por la reducción extrema de sus dedos, teniendo a penas 1 único dedo desarrollado en sus brazos. Su nombre significa "patas delgadas de Cerro Policía", tenía un tamaño muy pequeño de 60 cm de largo.
Gualicho shinyae (2016)
Terópodo de brazos extremadamente reducidos de la Formación Huincul (Río Negro) (~ 93 Ma). Descrito por Apesteguía y colegas; el nombre Gualicho es un espíritu maligno temido por los tehuelches y es en referencia a las complicaciones que sufrieron los investigadores para obtener los fósiles; el epíteto especifico honra a la técnica Shinya Inoue. Medía 6 m de largo y poseía brazos reducidos con solo 2 dedos en las cada mano. La reducción de brazos y dedos demuestra que era una convergencia a la que tendían varios grupos de dinosaurios carnívoros al aumentar de tamaño, como los abelisáuridos y los tiranosáuridos. Se sigue discutiendo a que grupo de terópodos se encontraría más cercanamente emparentado.
Overoraptor chimentoi (2020)
Paraviano de la Formación Huincul (Río Negro) del Cretácico superior (~ 97 - 93 Ma). Descrito por Motta y colaboradores. Su nombre significa "raptor overo de Chimento" en honor a el Dr. Roberto Nicolás Chimento quién descubrió los restos. Medía aproximadamente 1,3 m de largo con brazos y patas muy largas. Poseía una combinación mixta de características. Sus patas eran típicas de un raptor corredor con su clásica garra recurvada, sin embargo sus brazos tenían características anatómicas similares a las aves. Este fósil es importante para entender como fue la transición evolutiva entre raptores y aves.
SAUROPODA (herbívoros de cuello largo)
Grupo de dinosaurios herbívoros conocidos por ser los animales terrestres más grandes que han existido. Dominaron los ecosistemas terrestres desde el Triásico Superior (hace unos 210 millones de años) hasta su extinción al final del Cretácico (hace 66 millones de años).
REBBACHISAURIDAE
Son una familia dentro del grupo Diplocoidea que se caracterizan por tener largos cuellos y colas como látigos. En particular, los rebbachisauridae se distinguen de otros saurópodos por sus dientes característicos, que tienen un ángulo bajo, facetas de desgaste internas y esmalte asimétrico. Además de tener numerosos dientes de reemplazo rápido.
Zapalasaurus bonapartei (2006)
Rebbachisáurido de la Formación La Amarga (Neuquén) (Aptiano–Albiano ~ 130-122 Ma), descrito por Salgado y colaboradores. Estimado media 15 m de largo. Su nombre significa "reptil de Zapala de Bonaparte", Zapala es la localidad de procedencia y esta dedicado a el Dr. José Bonaparte. Poseía un cuello muy largo, que le permitía alimentarse a diferentes alturas sin tener que desplazarse demasiado.
Cathartesaura anaerobica (2005)
Rebbachisáurido de La Buitrera (Río Negro), Formación Huincul (97-93 Ma). De tamaño aproximado de 17 m de largo y 15 toneladas de peso, descrito por Gallina y Apesteguía. Cathartes es el género al que pertenecen los buitres, como este dinosaurio proviene de "La Buitrera" de ahí a que se llama "lagarto buitre". El epíteto específico viene por la empresa Anaeróbicos S.A. que proporciono apoyo logístico en la excavación. Presentaba un cuello largo y complexión ligera con un rango de movimiento dorso-ventral limitado.
Comahuesaurus windhauseni (2012)
Rebbachisáurido de la Formación Lohan Cura (Neuquén) (117 - 100 Ma), descrito por Carballido y colegas. Su nombre significa "reptil del Comahue de Windhausen", Comahue es la región en la que se lo encontró y en honor Anselmo Windhausen, quien contribuyó significativamente al estudio geológico de la Patagonia. Los restos fueron encontrados como "un lecho de huesos" que incluían 3 individuos distintos apilados. Su hallazgo ayuda a entender la evolución temprana de los rebaquisáuridos en América del Sur.
Lavocatisaurus agrioensis (2018)
Rebbachisáurido de la Formación Rayoso, Cuenca Neuquina (Aptiano–Albiano ~ 125 - 109 Ma), descrito por Canudo y colaboradores. El nombre genérico hace referencia a René Lavocat, un paleontólogo francés, mientras que el especifico a Agrio del medio que es la localidad donde se lo encontró. Cráneo notablemente completo, alargado y es muy similar a su pariente del norte, el Diplodocus. Su característica más notoria es la evidencia de la presencia de una cubierta queratinosa parecida a un pico en la parte anterior del hocico.
DIPLODOCIDAE
Fueron una familia de dinosaurios saurópodos que dominaron el paisaje durante el Jurásico Superior (hace unos 155-145 millones de años), principalmente en lo que hoy es Norteamérica, África y Europa.
Leinkupal laticauda (2014)
Diplodócido de la Formación Bajada Colorada (Cretácico temprano ~ 136,4 Ma) (Neuquén), descrito por Gallina y colaboradores. Su nombre viene del mapundungun Lein que significa "desvanecer" y kupal "familia" en referencia a que el último diplodócido conocido a escala global. El nombre de la especie viene del latín latus "amplio" y cauda "cola" porque presenta vertebras caudales anchas.
TITANOSAURIA
Los saurópodos de este grupo se caracterizan por tener cabezas pequeñas, dientes espatulados y forma corporal variada. Fueron el grupo más diverso y exitoso de saurópodos en el hemisferio sur durante el Cretácico. Se conocen no solo esqueletos sino también huellas, nidadas, y hasta coprolitos. En este grupo se encuentra Argentinosaurus (lognkosaurio),
Titanosaurios basales o de posición incierta
Representan formas tempranas o primitivas del grupo, a menudo conocidas por restos fragmentarios, principalmente del Cretácico inferior y medio. Muchos desarrollaron placas óseas (osteodermos) en el dorso para defensa.
Malarguesaurus florenciae (2008)
Titanosauriforme de la Formación Portezuelo, Malargüe (Mendoza) (Cretácico ~ 89 Ma). Descrito por González Riga y colaboradores. Se estima que media 18 m de largo y pesaba 20 toneladas. Su nombre significa "lagarto de Malargüe". Saurópodo robusto que se distingue por la forma única de sus vértebras caudales (de la cola), que presentan una morfología de transición entre grupos primitivos y los titanosaurios más avanzado.
Narambuenatitan palomoi (2011)
Titanosaurio de la Formación Anacleto (Neuquén) del período Cretácico (83 - 75 Ma), descrito por Filippi, García y Garrido. Su nombre significa "titán de Puesto Narambuena de Palomo" en referencia al lugar donde se lo encontró y a Salvador Palomo su descubridor. Se estima medía entre 12 y 14 m de largo.
Traukutitan eocaudata (2011)
Titanosaurio de la Formación Bajo de la Carpa (Neuquén), del Cretácico Superior (~ 85 Ma). Descrito por Juárez Valieri y Calvo. Su nombre Trauku es el nombre espíritu gigante de la montaña en la mitología araucana y la palabra titán. Eocaudata viene del griego eos que es "amanecer" y cauda que es "cola", en referencia a la forma primitiva ("basal") de sus vértebras de la cola. Caudales eran anfiplanas, poco comunes en titanosaurios. Se estima que medía unos 24 m de largo y pesaba aproximadamente 29 toneladas.
Notocolossus gonzalezparejasi (2016)
Titanosaurio gigante de la Formación Plottier (Mendoza) del Cretácico Superior (86 Ma), descrito por González Riga y colaboradores. Notocolossus proviene del griego notos que es "sur" y kolossos, "coloso". Y gonzalezparejasi rinde homenaje al Dr. Jorge González Parejas por su apoyo a las investigaciones paleontológicas en la región. Se estima media de 25 a 28 m de largo y pesaba entre 40-60 toneladas. El segundo espécimen preservó un pie trasero derecho completo, un hallazgo extremadamente raro en titanosaurios. Este pie muestra una estructura compacta adaptada para soportar un peso extraordinario.
Choconsaurus baileywillisi (2019)
Titanosaurio de la Formación Huincul, de Villa El Chocón (Neuquén), Cretácico superior (93 - 96 Ma). Descrito por Simón, Salgado y Calvo. Su nombre significa "lagarto de El Chocon de bailey willisi", en referencia al lugar del descubrimiento y en honor al geólogo estadounidense Bailey Willis quién realizó importantes estudios estratigráficos en la región a principios del siglo XX. Se estima midió 18 m de largo y pesaba 25 toneladas. Retiene rasgos anatómicos que sus parientes más evolucionados perdieron, como ciertos detalles en sus vértebras y manos.
Ninjatitan zapatai (2021)
Titanosaurio de la Formación Bajada Colorada (Neuquén) del Cretácico Inferior (140 Ma), descrito por Gallina, Canale y Carballido. El nombre Ninjatitan rinde homenaje al paleontólogo argentino Sebastián Apesteguía, apodado "el Ninja", mientras que zapatai a Rogelio Zapata, técnico del museo de Villa El Chocón que encontró el ejemplar. Se estima media 20 m de longitud. Considerado el titanosaurio más antiguo conocido; ancla el origen del clado cerca del inicio del Cretácico y refuerza la hipótesis de que este grupo se originó en Sudamérica.
Saltasauridos y afines (acorazados o de pequeño tamaño)
Se consideran los titanosaurios más evolucionados vivieron a finales del período Cretácico, hace aproximadamente entre 84 y 66 Ma. La mayoría eran relativamente pequeños para ser saurópodos, midiendo entre 8 y 15 m de largo. Fueron los primeros saurópodos en los que se confirmó la presencia de osteodermos (placas óseas en la piel), una armadura similar a la de los cocodrilos que les servía de defensa contra depredadores como los abelisáuridos.
Bonitasaura salgadoi (2004)
Titanosaurio de Cerro Policía (Río Negro), transición Bajo de la Carpa–Anacleto (85 Ma). Descrito por Apesteguía. El nombre es por la localidad del hallazgo, la cantera "La Bonita", mientras que salgadoi honra al reconocido paleontólogo argentino Leonardo Salgado. Medía unos 10 m de largo y pesaba alrededor de 5.5 a 6 toneladas. Su característica más inusual es su mandíbula cuadrada con un borde afilado detrás de los dientes frontales. Se cree que este borde estaba cubierto por un estuche de queratina (pico) que utilizaba para cortar vegetación dura. Caso de estudio en craneología y tafonomía patagónica.
Overosaurus paradasorum (2013)
Titanosaurio de Cerro Overo (Neuquén), Formación Bajo de la Carpa (Santoniano; 84-78 Ma). Descrito por Coria y colegas. Con una longitud estimada de solo 10 m y un peso de entre 5 y 7 toneladas. Su nombre significa "Lagarto de Overo", en referencia a la localidad donde fue descubierto, y paradasorum rinde homenaje a la familia Parada, quienes facilitaron el hallazgo y la excavación. El fósil se caracteriza por incluir una serie vertebral articulada excepcionalmente completa desde el cuello hasta la cola.
Sarmientosaurus musacchioi (2016)
Titanosaurio de Chubut (Bajo Barreal) del Cretácico (100-89 Ma), descrito por Martínez y colaboradores. Se estima media 12 m de largo y pesaba 10 toneladas. Sarmientosaurus hace referencia a la ciudad de Sarmiento, cercana al lugar del hallazgo; y musacchioi es un homenaje al Dr. Eduardo Musacchio, un geólogo y educador de la Universidad Nacional de la Patagonia San Juan Bosco. Tenía órbitas oculares muy grandes (40% de la longitud del cráneo), lo que sugiere una visión superior a la de otros dinosaurios similares. La estructura de su oído y cuello indica que habitualmente mantenía la cabeza inclinada hacia abajo, sugiriendo que se alimentaba principalmente de vegetación a nivel del suelo.
Nullotitan glaciaris (2019)
Titanosaurio de la Formación Chorrillo (Santa Cruz) del Cretácico (70 Ma), descrito por Novas y colegas. Se estima que alcanzaba entre 20 y 25 metros de largo. Nullotitan rinde homenaje al geólogo Francisco Nullo, quien descubrió los primeros restos en 1980. El epíteto glaciaris se debe a que el sitio del hallazgo ofrece una vista directa al Glaciar Perito Moreno.
Lognkosauria (los titanes de titanes)
Incluye a algunos de los animales terrestres más grandes y pesados que han existido, con la mayoría de sus restos hallados en Sudamérica, particularmente en la Patagonia argentina. Se caracterizaban por tener vértebras masivas con espinas neurales muy robustas y caja torácica extremadamente ancha. Aunque algunos miembros eran de tamaño "común" para un titanosaurio, otros representan los límites máximos de masa biológica terrestre. Vivieron durante el Cretácico Superior (entre los 101 y 66 Ma).
Mendozasaurus neguyelap (2003)
Lognkosaurio de Mendoza (Coniaciano; 89-86 Ma), descrito por González Riga. Su nombre proviene de la provincia de Mendoza y el término huarpe neguyelap, que significa "primer lagarto". Se estima que medía entre 18 y 22 m de largo, con un pedo de 16 toneladas. Poseía un cuello notablemente robusto con espinas neurales muy anchas y altas, además de contar con osteodermos (placas óseas en la piel) de forma esféricas para protección. Referente basal de Lognkosauria.
Puertasaurus reuili (2005)
Saurópodo de Formación Cerro Fortaleza Santa Cruz (Cretácico, 70 - 68 Ma), descrito por Novas y colaboradores. Nombrado en honor al técnico del Museo Edigio Feruglio, Pablo Puerta quien hizo el descubrimiento, y a Santiago Reuil quién reconstruyo la vertebra a partir de 60 fragmentos. Su vértebra dorsal (espalda) mide 1,68 m de ancho, lo que la convierte en la vértebra más ancha conocida de cualquier saurópodo. Solo se conocen 4 vértebras (una cervical, una dorsal y dos caudales), lo que es apenas el 3% de su esqueleto, dificultando estimaciones precisas de su tamaño total. Se cree que era tan grande como Patagotitan.
Futalognkosaurus dukei (2007)
Gigante lognkosaurio del Lago Barreales (Neuquén), Formación Portezuelo (Cretácico Superior, 87 Ma). Descrito por Calvo y colaboradores; se estima medía entre 26 y 30 m. y pesaba entre 30 y 50 toneladas. Su nombre proviene del mapudungun futa, "gigante", y lognko "jefe", sumado al griego saurus, "lagarto". Se traduce como "lagarto jefe gigante". El nombre de la especie, dukei es por la empresa Duke Energy, que patrocinó la excavación. Del ejemplar se recuperó alrededor del 70% del esqueleto, incluyendo un cuello casi completo y una pelvis inmensa de casi 3 m de ancho. Tenía 14 vértebras cervicales con espinas neurales muy altas en forma de "aleta de tiburón", lo que indica una musculatura extremadamente potente.

Patagotitan mayorum (2017)
Titanosaurio gigante del Miembro Cerro Castaño (Chubut), Cretácico (101 Ma). Descrito por Carballido, Pol, Cerda y colegas a partir de múltiples individuos. Fue hallado en 2012 por un peón rural en la estancia de la familia Mayo (de ahí el nombre mayorum). La excavación, liderada por equipos del MEF (Museo Edigio Feruglio) recuperó más de 150 huesos de al menos 6 individuos, lo que lo convierte en uno de los titanosaurios más completos conocidos. Se estima que medía unos 37 m de largo y pesaba 70 toneladas.

Chucarosaurus diripienda (2023)
Titanosaurio gigante de la Formación Huincul (Río Negro), Cretácico superior (90 Ma). Descripto por Agnolin y colaboradores. Se estima medía unos 30 m de largo y pesaba entre 40 y 50 toneladas. Su nombre proviene del quechua chúcaro, que significa "animal indomable" y del latín diripienda "revuelto o desordenado". A pesar de su colosal tamaño, poseía extremidades más esbeltas y gráciles en comparación con otros titanosaurios de dimensiones similares.
ORNITHISCHIA (ornitópodos y tireóforos)
Dinosaurios que vivieron desde el Triásico Superior hasta finales del Cretácico (de 228 a 66 millones de años).Eran predominantemente herbívoros u omnívoros oportunistas.
ORNITÓPODOS BASALES
Este es un grupo de dinosaurios corredores, ágiles y de tamaño pequeño a mediano, que parecen haber sido muy exitosos y diversos en la Patagonia.
Anabisetia saldiviai (2002)
Ornitópodo corredor de la Formación Cerro Lisandro (Neuquén), Cretácico Superior (95 a 92). Sus fósiles fueron hallados originalmente en 1985 por el agricultor local Roberto Saldivia Blanco, descrito por Coria y Calvo. El nombre es en honro a su descubridor y a la arqueóloga Ana María Biset. Era un herbívoro bípedo y ágil, de unos 2 m de largo y 20-25 kg de peso. Tenía extremidades traseras largas adaptadas para correr y una cola extensa para mantener el equilibrio. Se cree que vivía en ambientes pantanosos o zonas con abundantes ríos y vegetación densa
Talenkauen santacrucensis (2004)
Iguanodonto basal de la Formación Cerro Fortaleza (Santa Cruz), Cretácico Superior (76-70 Ma). Descrito por Novas y colaboradores. Su nombre significa "cráneo pequeño" en lengua Tehuelche, en referencia a las proporciones de su cabeza respecto a su cuerpo. Medía de 4 a 4.7 m de largo y pesaba unos 300 kg. Poseía placas óseas ovaladas a los lados de las costillas. A diferencia de una armadura, se cree que estas placas (procesos uncinados) ayudaban en la ventilación de los pulmones durante la respiración, de forma similar a las aves actuales. Era un herbívoro bípedo y ágil. Conservaba rasgos primitivos como dientes en la punta del pico y un primer dedo en el pie, características que sus parientes más evolucionados perdieron.
Isasicursor santacrucensis (2019)
Ornitópódo grácil de la Formación Chorrillo (Santa Cruz) Cretácico Superior (75-70 Ma), descrito por Novas y colaboradores. Su nombre significa "corredor de Isasi", en honor a su descubridor, el técnico paleontólogo Marcelo Pablo Isasi. Era un herbívoro bípedo de unos 4 a 5 m de largo y un peso estimado de 180 kg. Sus patas traseras estaban altamente adaptadas para la carrera, lo que sugiere que era un animal veloz. El hallazgo de individuos de distintas edades en un mismo sitio indica que probablemente vivían en manadas.
Mahuidacursor lipanglef (2019)
Ornitópodo pequeño, del Cerro Overo (Neuquén), Formación Bajo de la Carpa (Cretácico superior, 86-83 Ma). Descripto por Cruzado Caballero y sus colaboradores. Su nombre proviene de la palabra mapuche mahuida, que significa "montaña" o "cerro", y latí cursor, que significa "corredor". Y lipanglef significa "brazos ligeros". El nombre completo se traduce como "corredor de la montaña de brazos ligeros". Se estima media 6 m de largo. Era un dinosaurio herbívoro.
HADROSAUROIDEA
Son los ornitópodos más evolucionados, también conocidos como dinosaurios "pico de pato". Contaban con miles de dientes organizados en columnas complejas diseñadas para triturar vegetación dura antes de ingerirla.
Bonapartesaurus rionegrensis (2017)
Hadrosaurido de la Formación Allen (Río Negro) Cretácico superior, descrito por Cruzado‑Caballero y Powell. Su nombre rinde homenaje al paleontólogo argentino José Fernando Bonaparte. Se estima alcanzaba los 9 m de largo. Se caracteriza por tener espinas neurales extremadamente largas en sus vértebras sacras y caudales, con una altura que supera en más de 3 veces la del cuerpo vertebral. El holotipo destaca por presentar evidencia de enfermedades, incluyendo fracturas curadas en la cola y un posible tumor óseo (neoplasia) en su pie izquierdo, lo que ofrece datos únicos sobre la salud de estos dinosaurios.
THYREOPHORA
Este grupo incluye a los dinosaurios con armaduras dérmicas y defensivas. Eran exclusivamente herbívoros, con cabezas pequeñas y picos para procesar vegetación baja.
Jakapil kaniukura (2022)
Tireóforo basal de la parte alta de la Formación Candeleros (Río Negro), Cretácico Superior (97 - 94 Ma). Descrito por Riguetti, Apesteguía y Pereda‑Suberbiola. Medía 1,5 m de largo y pesaba entre 4 y 7 kg. Ja-Kapïl significa "portador de escudos" en puelche, y kaniukura significa "cresta de piedra" en mapudungun. Estaba protegido por hileras de osteodermos (huesos dérmicos) en forma de escudos que cubrían su cuello, lomo y cola. Era un bípedo herbívoro con una mandíbula fuerte y corta.
POSICIÓN INCIERTA
Isaberrysaura mollensis (2017)
Neornitisquio/tireóforo basal de la Formación Los Molles (Neuquén) Jurásico Medio (168-170 Ma), descrito por Salgado y colegas. El nombre hace referencia a la formación en la que se encontraba y a Isabel Valdivia Berry, quién es la que descubrió los restos. Se estima que medía entre 5 y 6 m, se cree que era mayormente bípedo y carecía de las placas óseas características de sus parientes más avanzados. El espécimen holotipo conservaba contenido estomacal fosilizado, consistente en una gran cantidad de semillas de cícadas. Esto sugiere que las tragaba enteras y actuaba como un importante dispersor de semillas en su ecosistema.
